
TRYING TO REASON WITH HURRICANE SEASON
As I write, river flooding and cleanup from Hurricane Florence in North and South Carolina are ongoing. The storm was not a major one in terms of maximum sustained winds--only a Category 1 on the Saffir-Simpson scale when it made landfall at Wrightsville Beach, near Cape Fear, NC. But the storm approached the coast very slowly, and moved only very slowly once it made landfall. That, and the areal extent of of the storm, resulted in quite a beating for the eastern Carolinas.

Satellite image of Florence approaching the Carolina coast.
The slow movement allowed the tropical cyclone to continue recharging its rain bands with warm, moist air from the Atlantic Ocean, resulting in 17 to 36 inches (432-914 mm) of rain in coastal N.C., and >10 inches (>250 mm) in many inland locations, in the middle and upper portions of the river basins. With respect to river flooding in the Carolinas, Florence draws comparisons to Hurricanes Floyd (1999) and Matthew (2016). The record peaks in most rivers of the have now occurred in one of those three events. But in the cases of Floyd and Matthew, the rain was falling on ground that was already wet and running off into streams that were already running high--in other words, the antecedent conditions made the flooding far more extensive than it would have been otherwise. Florence was different. As it approached the coast, conditions were relatively dry, and streams were at lower flow ranges (as they often are in September). Thus in terms of the hydroclimatological synoptic situation, Florence reminded me much more of Hurricane Harvey, which struck the Texas Coast in 2017.
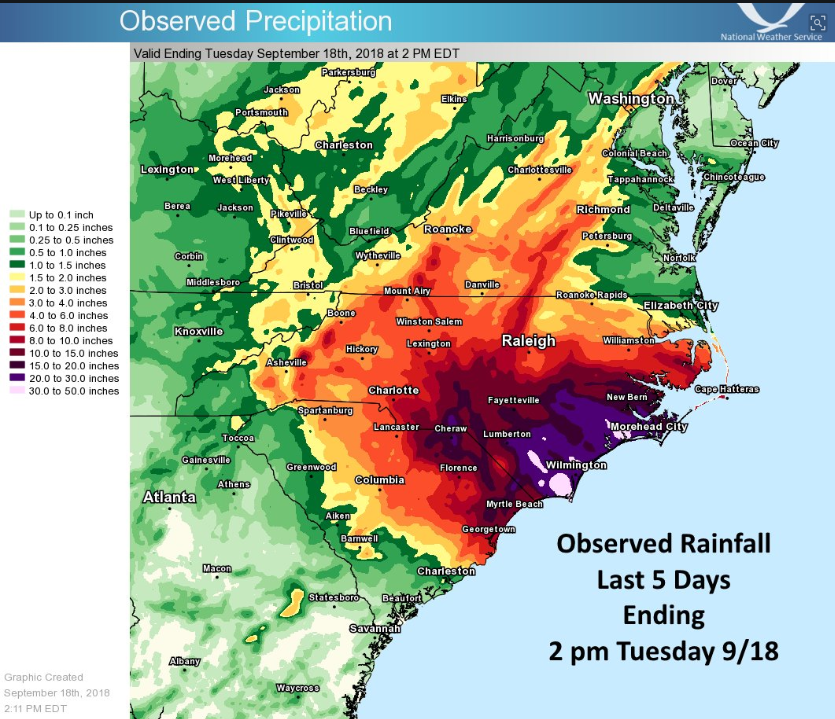
Rainfall from Florence (National Weather Service, Newport/Morehead City NC).
Harvey resulted in record storm rainfall totals for the USA; >50 inches (>1270 mm) in numerous locations, and massive rainfall flooding (in addition to wind, wave, and storm surge effects). While Harvey was a much larger storm than Florence in most respects, they had two things in common--very slow movement, and very large areal extent. The National Hurricane Center’s report on Harvey is here, and the U.S. Geological Survey’s study of flooding resulting from the storm is here.
A recent study showed that hurricanes have slowed their rate of movement by 10% in recent decades--and that doesn't include Harvey and Florence. The cyclone slowdown is consistent with a weakening in atmospheric circulation in the tropical parts of the planet, a result of global warming, study author James Kossin found. However, the mechanism behind the slowdowns are unclear.

Radar image at landfall. The storm moved only very slowly (about 8 km/hr) as it approached the coast and moved inland.
Regardless of what may be slowing down hurricanes, we can expect things to get worse (or better, if you like tropical cyclones) as the climate warms. It is well established that warm water is necessary for tropical cyclone development, and once formed, tropical storms are fueled by warmer water. Further, each 1 degree C increase in air temperature results in a 7 percent increase in saturation vapor pressure and how much water vapor the air can hold. This both provides more fuel for hurricanes (as release of latent heat as moisture condenses in the rising air of the low pressure center) and exacerbates the rain potential of a slow-moving storm.
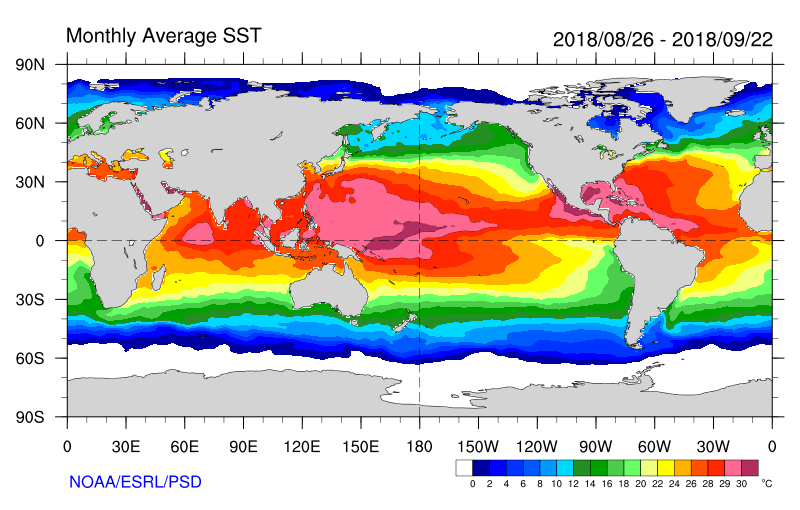
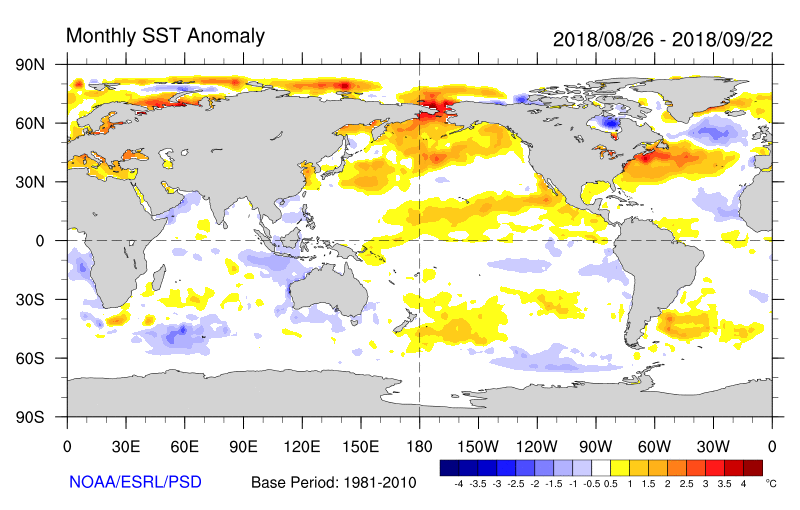
Sea surface temperatures, and temperature anomalies for the 28 August – 22 September period during which Florence and several other tropical cyclones formed. Note values in the North Atlantic and off the east coast of the U.S. Temperatures of 26 degrees C or greater facilitate tropical cyclone formation and strengthening.
Those of us interested in effects of contemporary and near-future climate change on coastal plain rivers have focused, appropriately enough, on sea-level effects. Recent events suggest, however, that perhaps climate warming induced changes in the frequency, severity, and behavior of tropical cyclones (and other storms?) may be of comparable importance. Mega-rainfall events such as Hurricanes Harvey and Florence not only send a lot of water down the channel, but they do so via precipitation concentrated in the lower portion of the drainage basins.

Cypress-Gum swamp near Croatan, NC covered with 0.5 m of sand during Florence. The pre-storm surface was organic muck.
Posted 27 September 2018































{kind=link}