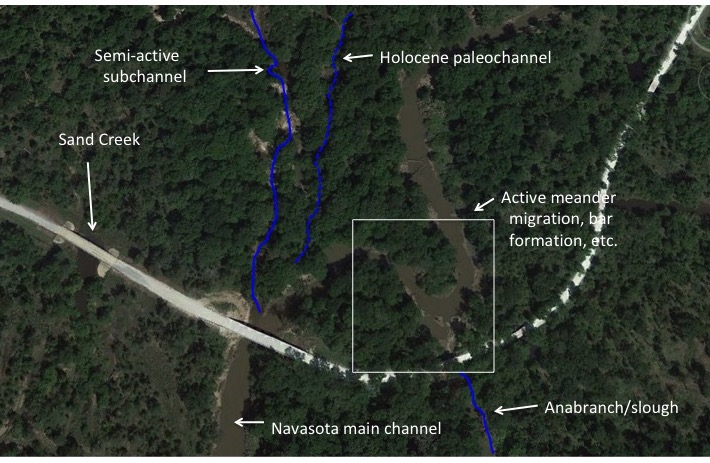
TRYING TO REASON WITH HURRICANE SEASON
As I write, river flooding and cleanup from Hurricane Florence in North and South Carolina are ongoing. The storm was not a major one in terms of maximum sustained winds--only a Category 1 on the Saffir-Simpson scale when it made landfall at Wrightsville Beach, near Cape Fear, NC. But the storm approached the coast very slowly, and moved only very slowly once it made landfall. That, and the areal extent of of the storm, resulted in quite a beating for the eastern Carolinas.

Satellite image of Florence approaching the Carolina coast.