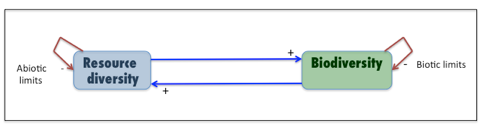
The Top 10 Forms of Complexity in Earth Surface Systems
When we (scientists) talk and write about complexity in recent years, the focus is on complex nonlinear dynamics, and related phenomena such as deterministic chaos, dynamical instability, some forms of self-organization, fractal geometry, etc. These are forms or sources of complexity that are intrinsic to the structure of dynamical systems, but these are hardly the only things that make systems complex. So, to make sure we don’t forget the elements of complexity that transcend so-called “complexity science,” I present the Top 10 Forms of Complexity in Earth Surface Systems (ESS). ESS is a blanket term that includes geomorphic systems, landscapes, ecosystems, soil systems, etc. Even though the items are numbered, they are actually in no particular order. Many ESS may exhibit only a few of these forms, and still be quite complex!
The list I was gonna do has already been done (http://grogsmovieblogs.com/).
Forms of Complexity in Earth Surface Systems