
Preferential flow occurs at all scales and in all phenomena in hydrological systems. By this we mean that rather than more-or-less uniform sheets of runoff moving across the ground surface, or more-or-less uniform bodies of water soaking into and seeping through soils and groundwater aquifers, most flow is concentrated in preferential flow paths.
Preferential flow of surface water—the universal tendency for flow to become channelized and form channel networks—is so obvious that it has been more or less taken for granted, though the mechanisms, and resulting structures and fluxes remain a highly active area of research. In soil and regolith, preferential flow has long been recognized, but its ubiquitous importance has only been widely recognized in recent decades. There are two main reasons for this, I think. Some forms of preferential flow, in soil pipes, large root channels, conduits, etc. are obvious, but other forms, such as macropore flow, are not readily recognized in the field. Second, a Darcian approximation of flow through porous media as a uniform mass often works fairly well, even where this type of flow is not necessarily dominant—presumably because the local preferential flow variations tend to average out in the aggregate.

An Ultisol in southwestern Louisiana. Like many soils, preferential flow paths are not necessarily evident at a cursory glance. Look more closely, however, and you seem numerous structural cracks, pin-hole to pencil-size pores, and roots & root channels.
But preferential soil moisture flux is ubiquitous, and may even occur (due to unstable wetting fronts) in relatively uniform sands. Preferential flow also occurs in various kinds of macropores and cracks, associated with, e.g., insect burrows, fine roots and root traces, interconnected soil pores, boundaries of soil aggregates and rock fragments, etc. For convenience I will use macropore to encompass all of these.

Sandy soil in coastal south Texas showing morphological evidence of preferential “fingered flow.”
In the last decade or so, substantial evidence has emerged that many soils (and groundwater systems) develop a dual-porosity structure, with slow flow and sometimes extensive storage in the matrix, and rapid flow with limited or transitory storage in macropores.
Henry Lin (2010) linked subsurface flow regimes and pedogenesis, pointing out that the dual partitioning of matrix and preferential paths goes a long way toward explaining the genesis of soil structure. He interpreted this in terms of the interplay of dissipating and organizing processes, and pointed out that the constructal theory of flux paths also predicts evolution toward flow partitioning and dual porosity.
Worthington (2019), working with groundwater systems, found that a high resistance matrix and low resistance preferential flow paths together form the minimum overall flow resistance; greater than that of a single-porosity system. There is in essence a division of hydrological labor, with the matrix taking care of storage and the preferential flow paths doing most of the transport work. The same phenomenon has also been interpreted in terms of coevolution of ecosystems, soils, and hydrology to satisfy contrasting hydrological functions of efficient drainage and sufficient storage of water and nutrients (Savenjie and Hrachowitz, 2017).
All of this makes perfect intuitive sense to me, is consistent with my field observations, and is supported by a great deal of empirical, model, and theoretical evidence. But I still have a bit of trouble wrapping my head around how it works at scales or conceptual levels in between the overarching principles of efficiency selection and organizing and dissipating processes, and the measurable particulars of a specific site or system.
Let’s consider the relationship between slow flow and soil moisture storage in the matrix, and rapid macropore flow. In any soil or hydrologic system, there is a given amount of available water in the form of excess precipitation. As a first approximation, partitioning this between matrix and macropores is a zero-sum game, and the two components can be considered competitive, with minus signs on the blue arrows in the figure below. That is, any H2O stored in the soil reduces the potential macropore flow, and vice-versa. The green arrows indicate self-effects; intrinsic constraints on the upper or lower values for negative self-effects, and positive feedbacks (self-reinforcement) for positive self-effects. Examples of some self-effects are shown in the table below.

|
Self-effects |
Matrix soil H2O storage |
Rapid macropore flow |
|
Positive |
Positive feedbacks between soil moisture & infiltration |
Positive feedbacks between flow & macropore or conduit size |
|
Negative |
Wilting point suction limits lower soil moisture; Field capacity limits maximum. |
Cross-sectional area, volume, conveyance capacity limit maximum fluxes |
With both self-effects negative, the system is dynamically unstable, indicating that a change will tip the system away from its pre-perturbation state—unless either component is near one of its limiting states, and these self-effects are stronger than the competitive relationship between the components. Suppose, for example, that soil moisture storage is approaching field capacity. Soil moisture is thus not competing with macropore flow, and increased macropore flow has little impact on soil moisture. The system tips to a saturated soil, macropore flow dominated state, allowing the excess moisture to drain away. This dynamically stable state exists until something (e.g., a change in the weather, rise of the water table to the surface) modifies the relationships (i.e., changes the sign of the green or blue arrows).
In another scenario we start with dry soil, with soil moisture at the wilting point—this negative self-effect dominates the system, as there is no macropore flow and no moisture to compete for. The beginning of a wetting event favors soil moisture, which wins the competition for water (limiting macropore flux), and may also switch to positive feedback as wetting fronts advance. This instability switches the system to a soil-wetting, limited macropore flux state. As (or if) soil moisture continues to increase, macropore flow comes into play and starts seriously competing with matrix storage, and stability is regained.
Yet another scenario is a super-wet state—the soil is saturated and maximum macropore flow is occurring, with the water table at or near the surface. Both self-effects are negative, but flow competition is irrelevant, creating a neutrally stable condition. After water input ceases, however, macropore flow begins to win the competition, with instability tipping the system to a wet soil, macropore dominated state. This facilitates more rapid drainage.
Other scenarios are possible, but these examples illustrate how the presence of both matrix storage and macropore flow allow the system to adjust to changing hydroclimatic conditions. Note that dynamical instability is necessary to allow the soil hydrological system to switch between different states.
But how, exactly, does the dual-porosity come into being?
Broadly speaking, it is a two-stage process. First, openings or macropores are created or the soil is segregated into high-resistance, tightly packed zones, and low-resistance potential flow pathways (PFP). Then preferential flow itself causes the dual porosity structure to persist (and in some cases become more pronounced).
The stage one segregation can occur due to a number of physical phenomena, including grain-packing, gravitational settling, shrink-swell and freeze-thaw processes, and boundaries or partings inherited from parent material. More important in many settings are biological processes, such as faunal tunneling and burrowing. Most important and interesting of all, to me, are the effects of plant roots, both exploiting, creating, and expanding PFPs.

Fig. 5 from Pawlik & Šamonil (2018): beech stump in a Haplic Cambisol, Turbacz, Gorce, Poland. a – general view, b – decaying roots and empty root channels, c – dense network of root channels
Persistence and potential growth of PFPs can occur due to mass transport and erosion (physical and chemical) along the PFPs. Certainly macropore deposition and clogging can occur, but the overall net effect is to maintain and sometimes enlarge the PFPs. With respect to biological features, fauna may maintain their tunnels. Plant roots both funnel infiltrated water along roots, and extract water via transpiration along these pathways. In addition, the roots and the associated microbes in the rhizosphere create chemical conditions that facilitate weathering and dissolved material transport. The pressure of root growth pushes soil material aside, packing it more tightly and reinforcing the high-flow-resistance matrix.
The specific details of these processes are highly variable, but one very recent and specific example examines changes in podzol morphology due to tree root modification of soil hydrology and porosity in a tropical rainforest in Brazil (Martinez et al., 2021). Pawlik and Šamonil (2018) showed how hydromorphic processes associated with tree roots persist in podzolized soils in three temperate forest settings, in ways consistent with development of dual porosity systems. Brantley et al. (2017) give an overview of how trees “build and plumb” the critical zone.
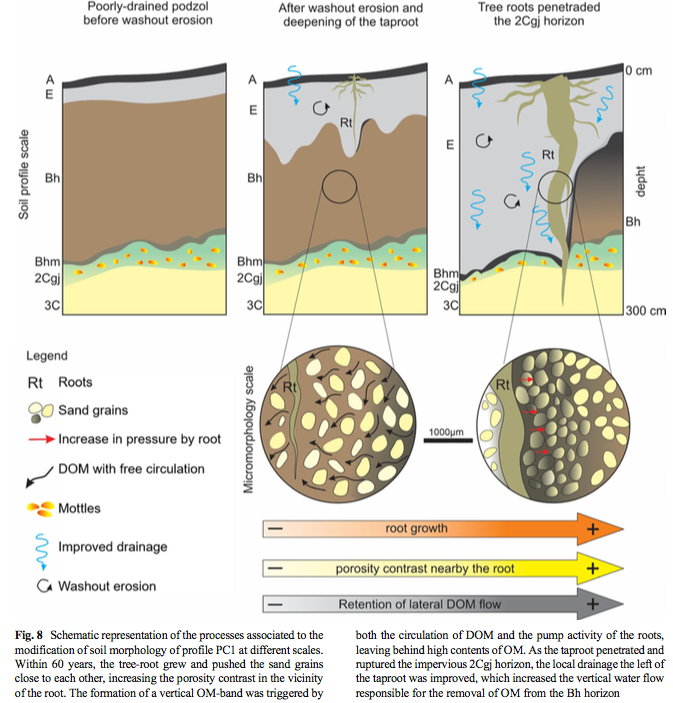
From Martinez et al., 2021
So plants, especially trees, help create dual-porosity soil and regoliths. This, in turn, provides a soil moisture storage reservoir for plants in the form of the matrix, and a mechanism for rapid drainage of excess water via the PFPs. This is, therefore, an excellent example of positive ecosystem engineering, and the coevolution of soil, hydrologic, and ecosystems.
References
Brantley SL, Eissenstat DM, Marshall JA, et al. 2017. Reviews and syntheses: on the roles trees play in building and plumbing the critical zone. Biogeosciences 14:5115–5142
Lin, H., 2010. Linking principles of soil formation and flow regimes. Journal of Hydrology 393: 3–19.
Martinez, P., Buurman, P., do Nascimento, D.L., et al. 2021. Substantial changes in podzol morphology after tree‐roots modify soil porosity and hydrology in a tropical coastal rainforest. Plant and Soil: https://doi.org/10.1007/s11104-021-04896-y.
Pawlik, L., Šamonil, P., 2018. Biomechanical and biochemical effects recorded in the tree root zone – soil memory, historical contingency and soil evolution under trees. Plant and Soil 426, 109-134.
Savenjie, H.H.G., Hrachowitz, M., 2017. HESS opinions: Catchments as meta-organisms—a new blueprint for hydrological modeling. Hydrology and Earth System Sciences 21: 1107-1116.
Worthington, S.R.H. 2019. How preferential flow delivers pre-event groundwater rapidly to streams. Hydrological Processes 33: 2373-2380.
Comments/questions: jdp@uky.edu